Różnorodne związki chemiczne są szeroko wykorzystywane przez mikroorganizmy, rośliny, a także zwierzęta do przekazywania informacji oraz do wywierania określonego wpływu na otoczenie, w procesie reprodukcji, zdobywaniu pożywienia, a także w celach obronnych poprzez wydzielanie substancji odstraszających, toksycznych, alarmowych, czy antybiotyków. Mogą to być również związki ułatwiające lub utrudniające koegzystencję. Są one niezmiernie istotne dla zachowania życia pojedynczych osobników i przetrwania gatunku. Substancje tego typu zostały nazwane związkami semiochemicznymi, inaczej związkami sygnalnymi (semio, z greckiego, oznacza sygnał). Wśród tych związków dużą grupę stanowią związki infochemiczne, a więc substancje służące do przekazywania informacji zarówno pomiędzy osobnikami tego samego gatunku, jak i z przeznaczeniem dla organizmów innych gatunków. Można by powiedzieć, że związki chemiczne zastępują w przekazywaniu informacji inne sygnały, jak na przykład wizualne, dotykowe czy dźwiękowe, gdyby to nie one były najbardziej pierwotnym sposobem porozumiewania się. Sygnały stosowane w przyrodzie do przekazywania informacji mają różne własności i swoistą charakterystykę. Z informacji zawartych w tab.1 wynika, że związki chemiczne, jako nośniki informacji, wykazują wiele korzystnych cech na tle innych sygnałów.
Tab. 1. Charakterystyka sygnałów nośników informacji.
|
Rodzaj sygnałów |
||||
|
Cechy sygnałów |
wizualne | dotykowe | dźwiękowe | chemiczne |
| Zasięg maksymalny | średni | b. krótki | daleki | b. daleki |
| Szybkość wymiany informacji | duża | duża | duża | średnia |
| Zdolność omijania przeszkód | słaba | słaba | wysoka | b. wysoka |
| Możliwość lokalizacji źródła | wysoka | wysoka | średnia | zmienna |
| Wydatek energetyczny | niski | niski | wysoki | niski |
| Chronologia pojawienia się | późno | wcześnie | późno | pierwotnie |
| Potencjalna pojemność informacji | duża | niska | duża | b. duża |
Feromony pośród związków infochemicznych zajmują szczególną pozycję i, jak dotychczas, zostały poddane najbardziej wnikliwym badaniom. Służą one do wywoływania zakodowanej reakcji fizjologicznej lub pożądanego zachowania u odbiorcy tego samego gatunku, w celu zdobycia pożywienia, obrony lub reprodukcji. Zarówno feromony, jak i związki allelochemiczne mogą wywoływać reakcję adaptacyjnie korzystną jedynie dla dawcy (+,-), tylko dla odbiorcy (-,+), lub dla obu organizmów (+,+). Natomiast podział związków allelochemicznych jest bardziej zdecydowany; noszą one odrębne nazwy w zależności od ukierunkowania korzyści.
Feromony były prawdopodobnie pierwszą formą komunikowania się najprostszych organizmów. Ich nazwa pochodzi z połączenia greckich słów pherein (przekazywać) i hormon (pobudzać). Główna różnica pomiędzy hormonami i feromonami, dwiema grupami bardzo ważnych związków biologicznie czynnych, polega na tym, że hormony są wytwarzane przez gruczoły lub tkanki w procesie wydzielania wewnętrznego z przeznaczeniem dla organizmu wytwarzającego, a feromony wydzielane na zewnątrz organizmu są adresowane głównie dla innego odbiorcy. Stosowanie feromonów stwierdzono, jak dotychczas, u ponad 1500 gatunków zwierząt (głównie owadów - 1100 gatunków), kilkudziesięciu roślin, a także szeregu bakterii i drożdży. Można jednak z dużą dozą prawdopodobieństwa przyjąć, że tym sposobem przekazywania informacji posługuje się większość organizmów żywych, ale wykrywanie feromonów jest niezmiernie trudne z uwagi na ich niskie stężenie nawet w obrębie źródła i brak specyficznych testów potwierdzających ich istnienie i działanie.
Wiele jest przykładów oddziaływania feromonów. Owady społeczne stosują feromony alarmowe wzywające do zespołowej obrony, inne feromony służą do zaznaczania ścieżek prowadzących do źródła pożywienia, znakowania terytorium polowania, miejsca zbiórki itp. Niektóre zwierzęta drapieżne wydzielają zapachy przyciągające ofiary, ale tych związków nie zalicza się do feromonów, lecz allomonów, jako że działają głównie na osobniki innego gatunku. Inne wabiki (atraktanty, feromony płciowe) są szeroko rozpowszechnione w świecie zwierzęcym. Feromony są wykorzystywane, między innymi, do przekazywania informacji na duże odległości, w nocy i poprzez terenowe przeszkody. Z dotychczasowych badań owadów wynika, że wypracowały one bardzo zróżnicowane sposoby wykorzystywania substancji chemicznych do przekazywania informacji, chociaż niektóre inne zwierzęta, np. gady czy ryby, również chętnie i szeroko korzystają z nich. Pierwszym odkrytym feromonem był bombykol, związek wydzielony z gruczołów samic jedwabnika, zdolny do zwabienia samców z odległości 11 km. O trudnościach związanych z izolacją i identyfikacją feromonów świadczą wysiłki zespołu A. Butenandta, który - podejrzewając ich istnienie - poszukiwał tych związków przez 30 lat, zanim w roku 1959 udało mu się wyizolować bombykol, określić jego rolę i miejsce wytwarzania. Do otrzymania kilkunastu mg tego związku zużyto 500 000 gruczołów wyizolowanych z samic jedwabnika. Poszukiwania feromonów jedwabnika Butenandt prowadził w Politechnice Gdańskiej.
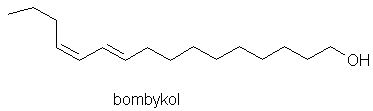
Rys. 1. Wzór bombykolu, feromonu płciowego jedwabnika
Wiele feromonów należy do związków organicznych o stosunkowo prostej budowie i niskiej masie cząsteczkowej, rzędu 80 - 300 D. Lotne feromony niskocząsteczkowe służą zwykle do szybkiego przekazywania informacji, ponieważ szybko ulegają rozproszeniu. Jeszcze krótszym czasem działania charakteryzują się cząsteczki feromonów zawierających nietrwałe grupy chemiczne, np. aldehydowe. Trwałe związki chemiczne, o większej masie cząsteczkowej, wykorzystywane są wówczas, gdy ich czas działania powinien być długi.
Feromony podzielono na 30 grup w zależności od pełnionych funkcji. Do najbardziej znanych należą feromony płciowe, ścieżkowe, agregacyjne, rozpraszające, alarmujące, stymulujące składanie jaj, dyskryminujące, zniechęcające, wprowadzające w błąd, czy znaczące. Czasami stosowane są feromony wielofunkcyjne, np. osy po użądleniu pozostawiają razem z jadem substancję alarmującą, a pszczoły wraz z jadem wydzielają śladowe ilości octanu izoamylu, znacznika wskazującego innym pszczołom miejsce do żądlenia. Kwas mrówkowy, typowy feromon obronny, może pełnić funkcję substancji alarmującej.
Do lat dziewięćdziesiątych uważano, że feromony są głównie preparatami jednoskładnikowymi. Odkrycie pierwszego feromonu, bombykolu, nadało na długi okres czasu niewłaściwy kierunek badaniom nad tymi związkami, ponieważ przyjęto jako sprawę oczywistą, że feromony są preparatami jednoskładnikowymi. To przekonanie było trudne do obalenia, ponieważ wiele składników feromonów występuje w stężeniach znacznie niższych niż składnik główny i ze względów technicznych były one trudne do wykrycia. Obserwowane różnice, często znaczne, aktywności głównego składnika (uważanego za właściwy feromon) w porównaniu z preparatem naturalnym, tłumaczono na różne sposoby, lecz nie brakiem dodatkowych składników. Bombykol okazał się nielicznym wyjątkiem, ponieważ preparat syntetyczny był równie aktywny jak feromon naturalny. Jednak ostatnie doniesienia wskazują, że nawet ten feromon zawiera śladowe domieszki, w tym bombykalu, aldehydu, który wydaje się mieć aktywność antagonistyczną (przeciwną). Obecnie wiadomo, że większość feromonów stanowi skomplikowaną mieszaninę, często z jednym dominującym składnikiem. Feromony mogą zawierać zarówno składniki lotne, jak i nielotne. Ich skład jest nie tylko gatunkowo zależny, ale zmienia się w obrębie tego samego gatunku w zależności od rodzaju przekazywanej informacji, pory roku, dnia, miejsca występowania gatunku, diety, przynależności do określonej kolonii, a nawet wieku osobnika. Autor osobiście doświadczył skutków regionalnego zróżnicowania feromonów, kupując w USA pułapki feromonowe na mrówki faraona. Okazały się one całkowicie nieaktywne w odniesieniu do mrówek faraona występujących w Polsce. W niektórych owadzich atraktantach płciowych doliczono się aż siedmiu aktywnych składników, przy czym obecność niektórych z nich jest konieczna do wywołania oczekiwanej reakcji u odbiorcy, a inne składniki modyfikują to zachowanie. Składniki tej mieszaniny mogą działać synergicznie (wzmacniająco) lub antagonistycznie. Ważny jest nie tylko ilościowy skład mieszaniny stanowiącej feromon, ale występowanie jego istotnych składników równocześnie. Muszą one w tym samym czasie dotrzeć do receptorów.
Świat naturalny jest bardzo zróżnicowany, dlatego też oprócz silnych tendencji do zachowania czystości gatunkowej znane są przykłady międzygatunkowych kontaktów pod wpływem identycznego feromonu płciowego. Już dawno zaobserwowano, że pomiędzy partnerami ciem rodziny Arctiidae, łatwo rozróżnialnych z powodu charakterystycznego ubarwienia, dochodzi do zalotów i kopulacji międzygatunkowej. Dopiero niedawno okazało się, że samice Callimorpha dominula L. i Tyria jacobaeae wydzielają ten sam atraktant płciowy. Prawdopodobnie gatunki te posiadają inny mechanizm zabezpieczający je przed hybrydyzacją.
Tab. 2. Sygnalizacja chemiczna zwierząt (tabela zaczerpnieta z Tablic Chemicznych Witolda Mizerskiego; opracowanie tabeli na potrzeby stron ChemFana: Witold Mozga - mozgaw@rocketmail.com).

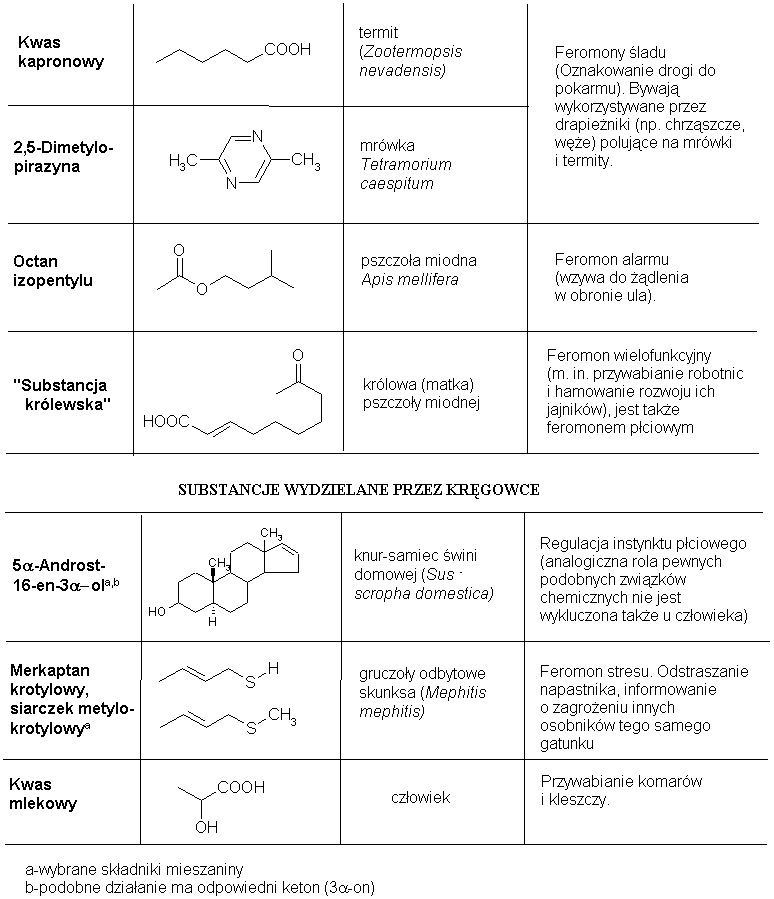
Identyfikacja
Prace nad identyfikacją feromonów zaczynają się od wydobycia ich z materiału biologicznego. W początkowym okresie badań nad feromonami izolowało się je wyłącznie z całych, homogenizowanych organizmów, ekstrahując skomplikowaną mieszaninę wielu związków. W trakcie oczyszczania następowały ogromne straty, co wymagało zabijania znacznej liczby osobników; jak pamiętamy, podczas izolowania bombykolu zabito 500 000 owadów. Obecny stan techniki pozwala wyekstrahować potrzebną ilość feromonu z gruczołów pobranych zaledwie od kilku osobników. Zostały również opracowane techniki pobierania feromonów bezpośrednio z gruczołów lub ze zbiorniczków, w których się znajdują. Identyfikacja składników feromonów za pomocą metod spektralnych wymaga potwierdzenia ich budowy poprzez porównanie związków naturalnych z substancjami otrzymanymi na drodze syntezy chemicznej. Związki syntetyczne stosowane są również zamiast produktów naturalnych do dalszych badań, a także do celów praktycznych, np. produkcji pułapek feromonowych. Istotnym elementem badania feromonów jest ocena ich aktywności biologicznej, a więc wykonanie testów oznaczania aktywności preparatu na każdym etapie, od pobierania materiału poprzez jego zagęszczanie i oczyszczanie, a w końcu w trakcie porównywania aktywności preparatów syntetycznych z naturalnymi. Nie ma ogólnych testów biologicznych, pozwalających na badanie aktywności wszystkich feromonów, zależą one od ich rodzaju i cech samych organizmów. Do najłatwiejszych należą testy na aktywność feromonów ścieżkowych, które polegają na naniesieniu badanej substancji na wąski pasek powierzchni i obserwowaniu reakcji owadów. W bardziej skomplikowanych badaniach stosuje się krzyżowanie szlaków. Nie sprawia też trudności badanie aktywności feromonów agregacyjnych, bo ich działanie można łatwo obserwować. Coraz częściej zamiast żywych owadów stosuje się ich części zawierające receptory feromonów. Najczęściej są to czułki, które - połączone przewodami ze wzmacniaczem - przekazują sygnał elektryczny do rejestratora. Tego typu testy noszą nazwę elektroantenografii, EAG (antena = czułki). Detektory te mogą być wykorzystywane wielokrotnie, a przez to, że reagują jedynie na związki wykazujące aktywność feromonów, są specyficzne dla określonego gatunku i niezwykle czułe, zdolne do rejestrowania aktywności naturalnej, a więc na poziomie kilku cząstek. Tak wysokiego progu wykrywalności nie zapewniają standardowe detektory. Nie są one jednak w stanie zastąpić organizmów żywych, tzn. wykazać ich prawdziwych reakcji na feromony. Odbierają one podobny sygnał zarówno od substancji agonistycznych, jak i antagonistycznych. Dla testowania małych ssaków stosuje się specjalne klatki, w których można obserwować reakcje zwierząt na badane substancje doprowadzane wraz z powietrzem.
Feromony produkowane są najczęściej w gruczołach zewnętrznych, np. owadzie, żeńskie atraktanty płciowe wytwarzane są często w gruczołach umieszczonych w membranach pomiędzy ostatnimi segmentami odwłoku, a samce wielkiej ćmy woskowej wytwarzają atraktanty w gruczołach umieszczonych w skrzydłach. Mrówka posiada około 50 zewnętrznych gruczołów produkujących związki semiochemiczne. Gruczoły te zostały podzielone na dwie grupy ze względu na rodzaj komórek, z których są zbudowane. Do jednej grupy należą gruczoły nabłonkowe produkujące i wydzielające substancje bezpośrednio do otaczającego je nabłonka. Do drugiego typu, bardziej zróżnicowanego, należą gruczoły zawierające wiele wyspecjalizowanych komórek wydzielniczych i kanałowych. Produkty tych gruczołów mogą być wydzielane bezpośrednio na zewnątrz lub gromadzone przejściowo w specjalnych zbiorniczkach. Gromadzony w zbiorniczku jad mrówek gatunku Formicidae, zawierający głównie kwas mrówkowy, którego stężenie dochodzi do 65 %, może stanowić aż 22 % wagi mrówki.
Oddziaływanie
Cząsteczki feromonu unoszone wiatrem w powietrzu, prądem wody, za pomocą dyfuzji lub w wyniku bezpośredniego kontaktu, docierają do odbiorcy. Do wywołania pożądanego efektu musi nastąpić ich kontakt z tą częścią ciała, w której znajdują się receptory. U owadów odbiornikiem feromonów lotnych są najczęściej czułki zawierające setki tysięcy komórek receptorowych. Zaadsorbowane na powierzchni czułek cząsteczki feromonu wnikają na zasadzie dyfuzji do ich wnętrza, gdzie poprzez system kanalików docierają do receptorów. Tam następuje ich rozpoznanie i przekształcenie chemiczne, czego rezultatem jest wytworzenie sygnału elektrycznego przesyłanego do mózgu. Mózg pod wpływem tego sygnału uruchamia skomplikowany proces, w tym syntezę lub uwalnianie enzymów, hormonów, czy przekaźników. Proces ten wywołuje specyficzne zachowanie odbiorcy zgodne z przesłaniem, którego treść była zakodowana w cząsteczce feromonu.
Czasami efekt działania feromonu zależy od jego stężenia. Jako przykład może służyć 4-metyloheptan-3-on, feromon mrówek Pogonomyrmex bodius, który przy niskim stężeniu zachęca robotnice do posłuszeństwa królowej, do wydajnej pracy, utrzymuje porządek w kolonii. Przy większym stężeniu wywołuje u robotnic agresję i atak, a pod wpływem jeszcze większego stężenia staje się dla nich poleceniem do kopania podziemnych korytarzy-kryjówek.
Częścią ciała ssaków, która reaguje na feromony, jest narząd przylemieszowy (vemoronosal organ, VNO). Do niedawna ten umocowany w przegrodzie nosowej podwójny, krótki, spłaszczony kanalik chrząstkowy (2-9 mm), wyścielony częściowo nabłonkiem migawkowym, był uważany za narząd zbędny dla człowieka, pozostałość pewnego etapu ewolucji. Ostatnio stwierdzono jednak, że dorośli ludzie też odbierają i reagują na feromony. U noworodków narząd przylemieszowy jest lepiej zachowany niż u dorosłych osób. VNO większości ssaków jest dobrze wykształcony i pełni ważną rolę odbiornika sygnałów chemicznych. Może być połączony nie tylko z nosem i służyć jako detektor substancji lotnych, ale poprzez połączenie z jamą ustną reaguje również na związki nielotne. VNO, uznawany za szósty zmysł, jest receptorem chemicznym funkcjonalnie i anatomicznie rozdzielonym od zmysłu węchu. Węch wykrywa substancje zapachowe, które mózg ocenia jako miłe, obojętne, nieprzyjemne, odrażające, czy nawet sygnalizujące niebezpieczeństwo. VNO reaguje natomiast na sygnały chemiczne, które aktywują stereotypowe, instynktowne zachowanie uruchamiane poprzez systemem neuroendokrynowy.
Feromony zwykle są aktywne w bardzo niskim stężeniu. Jako przykład służyć może wielkość aktywnego stężenia bombykolu wynosząca 10-21g/dm3, co odpowiada 2 500 cząsteczek w 1 cm3 powietrza. Dla porównania, w 1 cm3 powietrza znajduje się 2,7x1019 cząsteczek, głównie azotu i tlenu. Biorąc pod uwagę stężenie aktywne bombykolu, można oczekiwać wywołania określonego zachowania nawet pod wpływem pojedynczej cząsteczki feromonu. W praktyce wymagany jest kontakt z kilkoma cząsteczkami, co zabezpiecza przed przypadkowym uruchomieniem określonych reakcji. Bardzo aktywne są również feromony płciowe karaluchów - perifanon i supelapyron. Uwolnienie przez samice jedynie kilkuset cząsteczek tych związków uaktywnia samce nawet ze znacznej odległości. Fire ants reagują na swój feromon ścieżkowy już przy stężeniu 10 fg/cm ścieżki.
Feromony powierzchniowe zostały w literaturze wyodrębnione nie z uwagi na ich funkcję, jak w przypadku innych feromonów, ale ze względu na miejsce ich występowania. Są one zwykle trudno lotne, a więc działają z bliskiej odległości lub w wyniki bezpośredniego kontaktu, np. językiem, i służą do wzajemnego rozpoznawania się, prezentacji zajmowanej pozycji społecznej w grupie, przynależności kastowej, wywoływania określonych usług, np. iskania, mogą mieć też działanie afrodyzjaków. Ze skóry muszki owocówki (D. melanogaster), zarówno samic, jak i samców, wyodrębniono co najmniej 15 różnych węglowodorów, których część pełni rolę atraktantów. Stwierdzono wyraźne zróżnicowanie jakościowe i ilościowe składu mieszaniny węglowodorów w zależności od płci. Węglowodory te stymulują wstępną grę "miłosną" muszek, grają rolę swego rodzaju flirtu. Muszki pod wpływem feromonów zaczynają wykonywać specyficzne ruchy, przypominające taniec, a wibracja skrzydeł samców świadczy o ich płciowym pobudzeniu. Po kopulacji węglowodory pozostawione przez samca na powierzchni samicy obniżają jej atrakcyjność płciową i przez to nie budzi już ona zainteresowania innych samców. Węglowodory występujące w mniejszych stężeniach mogą wykazywać inne działanie, w tym synergiczne. Nieoczekiwane własności objawia wydzielany przez samców 7-pentakozen, który w dużych stężeniach wywołuje u nich skłonności homoseksualne.
Feromony powierzchniowe służą społecznym osom Polistes fuscatus do różnicowania robotnic najbliżej spokrewnionych, tj. sióstr, czyli córek jednej królowej, i dalszych krewnych, np. kuzynek, tj. robotnic wyklutych z jaj siostry królowej, oraz od niespokrewnionych osobników tego samego gatunku. Głównymi składnikami heksanowego ekstraktu powierzchni ciała os okazały się węglowodory, z których znaczna część służy do rozpoznawania stopnia pokrewieństwa. Skutkiem rozpoznania może być akceptacja lub odrzucenie robotnicy przez dominujące w kolonii ''ciotki''. Skład i udział poszczególnych węglowodorów w feromonie os rozróżniającym stopień pokrewieństwa jest zmienny i dostosowany do danej rodziny czy kolonii.
Rozprzestrzenianie
Szybkość rozchodzenia się związków chemicznych w powietrzu można ująć wzorem matematycznym. Wzór taki wyprowadzony na podstawie prawa dyfuzji, uwzględniający lotność feromonu, pozwala obliczyć czas, po jakim wymagane stężenie substancji znajdzie się w określonej odległości od źródła. Posługując się tymi zależnościami obliczono minimalne stężenia feromonów potrzebne do wywołania właściwej reakcji. Dla niektórych feromonów ścieżkowych promień działania wynosi 0.6 cm, a pewnych feromonów płciowych 600 m i więcej. Własności poszczególnych feromonów zależą od ich przeznaczenia, np. ścieżka zaznaczona feromonem ścieżkowym powinna być wąska, a czas jego aktywności stosunkowo długi, rzędu godzin. Natomiast feromon alarmowy powinien działać szybko, ale krótko. Nie może on pozostawać długo w powietrzu, żeby nie utrzymywać kolonii niepotrzebnie przez dłuższy czas w stanie gotowości bojowej. Promień działania feromonu alarmowego pewnego gatunku mrówek wynosi 10 cm, a po 2 minutach jego stężenie spada poniżej stężenia aktywnego. Feromony działające w powietrzu charakteryzują się dużą lotnością, natomiast dla feromonów rozchodzących się w wodzie ważnym czynnikiem jest hydrofilowość; wśród nich spotyka się aminokwasy, a nawet białka. W środowisku wodnym szybkość rozchodzenia się feromonów jest znacznie mniejsza niż w powietrzu, natomiast czas zaniku jest wielokrotnie dłuższy.
Selektywność
Selektywność feromonów zależy od ich budowy i pełnionej funkcji. Przykładem feromonów mało specyficznych są cząsteczki 2-heptanonu i octanu butylu różniące się znacznie własnościami chemicznymi. Mają one jednak zbliżony kształt oraz zawierają po jednej grupie karbonylowej i jako feromony mogą wywoływać podobne reakcje.
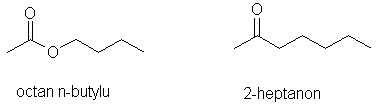
Rys. 2. Przykłady feromonów niespecyficznych
Większość receptorów feromonowych jest w stanie rozróżnić cząsteczki poszczególnych izomerów, w tym stereoizomerów. Szczególnie wysokie wymagania specyficzności natura postawiła feromonom płciowym. W ich przypadku zróżnicowanie dotyczy nie tylko identyczności chemicznej cząsteczki, jej konfiguracji, ale często są to skomplikowane mieszaniny, których skład jest uzależniony gatunkowo i regionalnie, może być inny nawet dla osobników sąsiadujących kolonii.
Rola konfiguracji
Duża część cząsteczek pełniących rolę feromonów zawiera centra chiralne i zwykle tylko jeden stereoizomer jest biologicznie czynny. Bombykol, pierwszy odkryty feromon, ma dwa podwójne wiązania, a wobec tego może występować w postaci czterech stereoizomerów. Feromonem jedwabnika jest izomer 10E,12Z. W literaturze nie ma danych na temat aktywności biologicznej pozostałych jego izomerów. Dużo natomiast uwagi poświęcono poznaniu zależności aktywności biologicznej cząsteczek feromonów od konfiguracji centrów chiralnych. Dotychczas zbadano aktywność ponad 200 stereoizomerycznych feromonów. Zaproponowano pogrupowanie ich w zbiory o określonych zależnościach aktywności biologicznej od konfiguracji. Do typu I należą te feromony, których tylko jeden z enancjomerów jest biologicznie aktywny, a drugi nie przejawia aktywności feromonu w stosunku do badanego organizmu. Jest to najczęściej spotykana zależność, obejmuje ona 60 % zbadanych związków. W typie II aktywność jednego stereoizomeru jest inhibowana przez inny stereoizomer. Do typu III zaliczane są feromony, w których nieaktywny lub mniej aktywny stereoizomer działa synergicznie na główny składnik feromonu, tak że ich mieszanina jest aktywniejsza od pojedynczego składnika. Znajomość tych zależności ma nie tylko znakomitą wartość naukową, ale również praktyczną, np. przy produkcji pułapek zawierających feromony. Nawet śladowa zawartość inhibitora może znacznie obniżyć aktywność pułapek. Pamiętać należy, że produkty przemysłowe rzadko osiągają czystość powyżej 95 %, a odczynniki chemiczne o czystości 98 % są uważane za czyste. Usuwanie śladowych zanieczyszczeń innymi związkami jest czasochłonną i kosztowną procedurą. Jeszcze trudniej otrzymuje się związki o wysokiej czystości chiralnej.
Wybrane grupy feromonów
Feromony płciowe są związkami chemicznymi wykorzystywanymi w procesie nawiązywania z partnerem wstępnej gry prowadzącej do parzenia się i zapłodnienia. Feromonami płciowymi mogą być różne związki chemiczne, w tym kwasy karboksylowe, ich estry, ketokwasy, aldehydy, fenole, alkohole, węglowodory, związki aromatyczne i alifatyczne. Często przedstawiciele przeciwnej płci żyją osobno, niejednokrotnie okres płodny samic jest krótki; to utrudnia procesy kojarzenia. Feromony mają na celu pokonywanie tych utrudnień poprzez kojarzenie partnerów, synchronizację czynności prowadzących do zapłodnienia. Feromony płciowe są na tle innych feromonów dobrze poznane, mimo to dla wielu gatunków nadal stanowią zagadkę. Aktywność seksualna mrówek jest limitowana do kilku dni w ciągu roku. Ograniczenie to oraz fakt, że wiele gatunków mrówek parzy się w czasie lotu, utrudnia prowadzenie badań ich życia seksualnego. Może dlatego nie udało się, jak dotychczas, zidentyfikować typowych feromonów płciowych u mrówek, chociaż stwierdzono ich istnienie. Znane są mrówki, których samice pozbawione skrzydeł przebywają blisko gniazda na ziemi lub roślinach i do nich zlatują się skrzydlate samce. Należy przyjąć, że samice przywabiają je do siebie lotnymi feromonami płciowymi. Przykładem wykorzystywania innych feromonów w procesie rozmnażania mogą być mrówki, których samce z różnych kolonii gromadzą się w jednym miejscu tworząc rój, do którego zlatują się samice. W tym przypadku rolę wabika odgrywa feromon agregacyjny. Zaś samce innego gatunku Megaponera foetus podążają do gniazda ścieżką zaznaczoną przez robotnice i do parzenia dochodzi wewnątrz gniazda. Tym razem funkcję wabika spełniają feromony ścieżkowe. Feromony płciowe są bardzo istotne dla owadów, zwierząt wodnych, węży, a także ssaków. Rozmnażanie płciowe mikroorganizmów również zaczyna się od wydzielenia feromonów.
Feromonami ścieżkowymi znaczony jest wąski pasek w terenie, który łatwo rozpoznają i mogą wzdłuż niego podążać osobniki tworzące wspólnotę. Tego typu chemiczny sposób znaczenia ścieżek stosują często termity, mrówki, czy gąsienice. Ścieżki takie prowadzą do źródła pożywienia lub miejsca założenia kolonii. Stwierdzono, że niektóre gatunki bezżądłowych pszczół stosują substancje chemiczne nawet do znakowania korytarzy powietrznych. O chemicznej naturze znakowania ścieżek przez mrówki wiedziano już od drugiej połowy XVIII w. Przez długi okres sądzono, że robotnice zostawiają na drodze swój zapach automatycznie przez stawianie stóp. Dopiero w XX w. udowodniono, że feromony ścieżkowe są wydzieliną z odwłoka, celowo rozsmarowywaną przez osobnika mającego za zadanie zaznaczyć drogę. Mrówki znaczą ścieżkę w drodze powrotnej do gniazda, po znalezieniu źródła pożywienia. Z tego wynika, że robotnice, które po znalezieniu źródła pożywienia powracają do gniazda jako pierwsze, nie korzystają ze ścieżki. Muszą więc mieć inny sposób orientacji w terenie i zapamiętywania drogi powrotnej. Feromony ścieżkowe często są złożoną mieszaniną. Ich skład zależy nie tylko od gatunku, ale również od rodzaju pożywienia, jego zasobów, a także od przynależności do rodziny, podrodziny, czy nawet kolonii. Feromony te są produkowane przez różne gruczoły. W trakcie izolacji i identyfikacji pierwszego feromonu ścieżkowego, wydzielanego przez mrówki Atta texana, poddano ekstrakcji 3,7 kg mrówek.
Znane są zwierzęta reagujące na feromony ścieżkowe innych gatunków. Korzystają one z dóbr odkrytych nie przez siebie.
Feromony znaczące terytorium są stosowane do oznakowywania granic miejsca zamieszkiwania lub polowania. Stosowane są one szeroko nie tylko przez owady, ale i przez inne zwierzęta, w tym ssaki drapieżne, które substancjami zawartymi w moczu zaznaczają granice swoich terenów łowczych. Pośród owadzich feromonów znaczących odkryto węglowodory alifatyczne i terpenoidy.
Feromony alarmowe wydzielane w wypadku zagrożenia kolonii, czy gniazda, wywołują całą gamę zachowań u odbiorcy. Reakcją na feromon alarmowy może być ucieczka, bezruch, szukanie schronienia, agresja. Rolę feromonów alarmowych pełnią alifatyczne i cykliczne alkohole, aldehydy, ketony, kwasy, estry, węglowodory, heterocykle, sulfidy, terpenoidy i inne.
Feromony agregacyjne powodują gromadzenie się osobników jednego gatunku w miejscu, w którym ten feromon został wydzielony przez przedstawiciela lub przedstawicieli tego samego gatunku w celu zwabienia partnerów seksualnych, przekazania informacji o znalezionym obfitym źródle żywności, czy też o odkryciu miejsca do założenia kolonii. Feromony tego typu mają stosunkowo długą trwałość. Undekan, wydzielany przez robotnice jednego gatunku mrówek, zachowuje aktywność przez 12 godzin.
Wyizolowanie feromonu agregacyjnego owada Cosmopolites sordidus daje nadzieję na opracowanie skutecznych metod ograniczenia populacji głównego szkodnika bananowców. Jego larwy żywią się kłączami tych roślin. Istotnym składnikiem tego feromonu jest (+)-sordydyna.
Feromony rozpraszające mają za zadanie utrzymywanie osobników tego samego gatunku w określonej odległości od siebie w celu ograniczenia rywalizacji wewnątrzgatunkowej, w tym walki o żywność, partnerów płciowych, miejsca składania jaj, zakładania gniazd itp.
Feromony dyskryminujące powodują, najczęściej we wczesnym stadium rozwoju, hierarchiczny (kastowy) podział osobników. Tego typu podział stosują owady społeczne, np. pszczoły, mrówki i termity. Samice pszczół mogą być robotnicami stanowiącymi zdecydowaną większość kolonii lub królową-matką. Ten podział, będący swego rodzaju dyskryminacją, odbywa się pod wpływem mieszaniny ośmio- i dziesięciowęglowych kwasów karboksylowych. Mieszanina kwasów utrzymująca status robotnic nie jest typowym feromonem, ponieważ jest wytwarzana przez ich gruczoły szczękowe. Pełni jednak rolę feromonu, gdyż służy jako dodatek konserwujący pożywienie larw, a przede wszystkim powstrzymuje rozwój larw żeńskich w kierunku królowej. Kwasy karboksylowe wytwarzane przez królową wchodzą w skład szczękowego feromonu królowej (QMP). Są one silnymi atraktantami wymuszającymi posłuszeństwo wobec królowej, udzielanie jej pomocy, tworzenie orszaku wokół królowej, skłaniają do pracy. Praca robotnic polega przede wszystkim na opiece nad jajami i larwami, ochronie i obronie gniazda oraz zdobywaniu pożywienia. Głównym zadaniem królowej jest reprodukcja, czyli składanie jaj oraz przywodzenie kolonii. Posłuszeństwo członków kolonii osiąga za pomocą wyżej wspomnianego feromonu QMP. Feromon ten tłumi zdolność robotnic do reprodukcji. Po 2-3 tygodniach bezpłodności królowej, lub z powodu jej braku, robotnice uzyskują zdolność składania jaj. Ta z robotnic, która pierwsza osiągnie tę zdolność, zaczyna wydzielać QMP i tym samym powstrzymuje transformację innych robotnic w kierunku królowej. W gruczołach robotnic i królowej znaleziono śladowe ilości składników feromonu o przeciwnym działaniu, co świadczy, że ich gruczoły zachowują zdolność produkcji obu feromonów. Wytwarzane są z tego samego substratu, jakim jest kwas stearynowy. U mrówek Solenopsis invicta zapłodniona królowa produkuje feromon uznający królową, który czyni ją atrakcyjną dla robotnic i motywuje je do służby na rzecz królowej. Aktywnym składnikiem tego feromonu jest mieszanina nienasyconego cyklicznego ketonu i zbliżonego pod względem budowy laktonu.
Feromony obronne i jady wytwarzane i wydzielane w celach obronnych lub ułatwiających zdobywanie pożywienia są związkami wywołującymi nieprzyjemne odczucie, ból, porażenie, a nawet śmierć. Związki te, kiedy nie są przeznaczone dla osobników tego samego gatunku, nie są zaliczane do klasycznych feromonów. Dobrze znany jest jad obronny mrówek. Skład tego jadu jest zróżnicowany. Głównym składnikiem jadu mrówek Formicidae jest kwas mrówkowy. Oprócz kwasu mrówkowego, w jadzie tym znajdują się krótkie peptydy i wolne aminokwasy w stężeniu nie przekraczającym 5 %. Jad mrówek nie należących do Formicidae zawiera dużo białek - enzymów, wśród nich fosfolipazy i hialuronidazę. Niektóre gatunki mrówek, np. myrmicine, produkują alkaloidy. Robotnica S. geminata może zgromadzić w swoim ciele 19 mg alkaloidów. Jad fire ants, wywołujący miejscowe zapalenie w okolicy użądlenia, zawiera 2-alkilo-6-piperydyny. Ćmy Utetheisa ornatrix, ich jaja oraz larwy są niejadalne dla większości zwierząt, ponieważ zawierają alkaloid odstraszający potencjalnych amatorów. Głównymi składnikami jadów węży są peptydowe i białkowe enzymy.
Rola feromonów w życiu ssaków
Jak dotychczas rolę feromonów, ich struktury czy działanie poznano najlepiej w świecie owadów. Już od dawna podejrzewano stosowanie feromonów przez ssaki. Dopiero niedawno nauka dostarczyła dowody na ich występowanie u ssaków, w tym zwierząt hodowlanych. Co spowodowało tak znacznie opóźnienie w poznaniu feromonów ssaków? Otóż ich izolacja, identyfikacja, analiza aktywności napotyka u ssaków na wiele trudności, związanych nie tylko z niskim poziomem stężeń. Znacznie trudniej jest znaleźć dla ssaków testy pozwalające na badanie ich reakcji na feromony. Nie można dla nich stosować elektroantenografii, powszechnie używanej w badaniu owadów. Jako przykład trudności napotykanych podczas badania feromonów ssaków mogą służyć wysiłki zmierzające do poznania feromonu płciowego wydzielanego przez krowy w okresie płodnym. Z najnowszych badań wynika, że feromon ten znajduje się nie w wydzielinie z pochwy, jak do niedawna przypuszczano, ale w moczu. Udało się nawet wydzielić frakcję aktywną, oznaczyć granice masy cząsteczkowej aktywnego związku (50 - 130 D) oraz określić jego podstawowe własności. Do 1997 r. nie udało się jednak wyizolować tej substancji w stanie czystym i w takiej ilości, żeby można było ją zidentyfikować. Badania feromonów w życiu ssaków prowadzi się poprzez obserwacje ich reakcji i zachowania. Stwierdzono między innymi, że przebywanie dorosłych samców pośród samic tego samego gatunku, np. myszy, owiec, kóz, bydła, świń czy koni, wpływa na rozwój płciowy młodych samic, przyspiesza rozpoczęcie funkcjonowania ich jajników oraz reguluje częstotliwość występowania rui. Substancje aktywne wywierające to działanie znajdują się w moczu samców i sam mocz dojrzałych płciowo samców wykazuje podobną aktywność. Nieaktywny jest mocz kastratów, a także samców niedojrzałych płciowo i starych. Ostatnie badania wykazały, że głównym składnikiem tych feromonów są androgeny. Samice z kolei wydzielają w czasie rui substancje sygnalizujące ich stan. Stymulują one organizmy samców do wytwarzania androgenów, a tym samym wzmagają ich aktywność seksualną. Mocz samic myszy ma inne działanie na samice przebywające w tej samej grupie. Spowalnia dojrzewanie płciowe młodych samic oraz zaburza rytmiczność rui. U szczurów feromony preowulacyjne skracają cykl menstruacyjny towarzyszących samic, podczas gdy feromony postowulacyjne wydłużają ten okres. Ostatnie doniesienia mówią o wykryciu podobnych zależności u kobiet.
Feromony lotne odgrywają u ssaków dużą rolę w trakcie nawiązania kontaktu między partnerami. Dużo zwierząt żyjących w niewoli zatraca zdolności reagowania na te feromony, ich szósty zmysł ulega stępieniu, co stanowi główną przyczynę trudności w rozmnażaniu niektórych zwierząt przetrzymywanych w niewoli, np. w ZOO. Są jednak gatunki, którym niewola nie przeszkadza w rozmnażaniu się. Należą do nich przede wszystkim gryzonie, a także zwierzęta udomowione. Szczególnie wdzięcznym obiektem w badaniach feromonów płciowych są chomiki. Już ponad 20 lat temu stwierdzono, że samiec chomika złocistego żywo reaguje na zapach wydzieliny pochwy samicy będącej w rui. Dla ułatwienia prowadzenia tych doświadczeń zbudowano izolowane od otoczenia klatki, do których można było w sposób selektywny doprowadzać wraz z powietrzem lotne substancje i obserwować reakcje zamkniętych w nich samców. System filtrów, pomp i urządzeń dozujących pozwala na dostarczanie do klatki powietrza czystego lub zawierającego określone związki chemiczne i usuwanie powietrza zużytego. Chomiki reagują zwykle bardzo szybko na feromony płciowe - już po 3 minutach kierowały się do otworu, przez który było doprowadzane powietrze z domieszkami i prezentowały zespół aktywności seksualnej. Wydzielina pochwy samicy chomika w okresie płodnym jest skomplikowaną mieszaniną wielu związków. Pośród substancji lotnych najbardziej aktywnym okazał się disiarczek dimetylu, którego aktywność stanowi jednak jedynie 20-40% aktywności lotnej frakcji próbki naturalnej. Aktywność feromonu płciowego wykazuje również frakcja nielotna, z której w 1996 r. wydzielono szereg rozpuszczalnych w wodzie białek o wysokiej aktywności afrodyzjakalnej. Z obecnego stanu badań wynika, że lotne feromony płciowe wydzielane przez samicę chomika wywołują u samca zainteresowanie samicą, a dopiero w wyniku bezpośredniego kontaktu z nielotnymi białkami uzyskuje on pełną aktywność kopulacyjną. Detektorem aktywnych białek jest narząd przylemieszowy (VNO). Odkrycie nielotnego afrodyzjaka znajdującego się w wydzielinie z pochwy tłumaczy zachowanie samców wielu gatunków ssaków obwąchujących żeńskie narządy płciowe przed kopulacją.
Niezmiernie istotną rolę pełnią feromony płciowe w rozmnażaniu słoni, ponieważ w naturze dorosłe osobniki płci przeciwnej żyją oddzielnie. Do ich kojarzenia dochodzi pod wpływem lotnego feromonu wydzielanego z moczem przez samicę słonia azjatyckiego kilka tygodni przed osiągnięciem okresu płodnego. Feromon ten wywołuje łatwo widoczną, charakterystyczną reakcję samca, zaczynającą się od specyficznego ruchu trąbą ułatwiającego dotarcie feromonu do VNO. Po pewnym czasie samiec zaczyna poszukiwanie samicy wydzielającej feromony. Tak wygląda początek godów słoni azjatyckich, trwających kilkanaście tygodni. Bardzo charakterystyczny ruch trąbą umożliwia badanie reakcji na działanie feromonów przez samce słoni trzymanych w niewoli; jest swoistym testem, który ułatwił postęp prac związanych rozpoznaniem feromonu płciowego słoni. Prace nad izolacją i identyfikacją głównego składnika feromonu płciowego słoni azjatyckich zajęły 15 lat i zakończyły się sukcesem w 1997 r. Tym związkiem okazał się octan Z-7-dodeceno-1-ylu. W naturalnej próbce stwierdzono występowanie również izomeru E na poziomie 3%. Aktywność biologiczna czystego octanu izomeru Z i z dodatkiem izomeru E jest nadal niższa od próbki naturalnej, co sugeruje, że naturalny feromon jest bardziej skomplikowaną mieszaniną. Odkrywcy feromonu płciowego słoni azjatyckich podjęli ostatnio prace nad wykryciem feromonów płciowych słoni afrykańskich.
Identyfikacja feromonu płciowego słoni azjatyckich stała się pewnego rodzaju sensacją naukową, nie tylko z tego powodu, że, jak dotychczas, znane są feromony płciowe jedynie kilku ssaków, ale okazało się, że główny składnik feromonu płciowego słonia jest identyczny z głównymi składnikami feromonów płciowych ponad 120 gatunków owadów, głównie Lepidoptera. Obecnie nie sposób jeszcze wytłumaczyć co spowodowało, że tak różne zwierzęta, jak słoń azjatycki i wiele owadów wykorzystują ten sam związek jako feromon płciowy.
Dużym zainteresowaniem cieszą się badania feromonów zwierząt hodowlanych. Stwierdzono, między innymi, że u świń maciora przyjmuje pozycję wygodną do kopulacji pod wpływem szeregu metabolitów testosteronu zawartego w ślinie knura. Pośród tych związków największą aktywność wykazuje 5<F128>a<F255>-androsteron. Ma on tak silne działanie, że został nazwany deprawatorem. U owiec wieloskładnikowy feromon płciowy znajduje się w tłuszczu pokrywającym wełnę tryka. Natomiast wydzielina gruczołów tłuszczowych samca bobra zawiera co najmniej osiem składników, przy czym sześć z nich musi być obecna, żeby wywołać reakcję prokopulacyjną u samic.
Powszechnie znana jest reakcja psów na obecność suki w okresie cieczki. Psy wyczuwają jej stan z dużej odległości. Jest to także wynik działania lotnych feromonów rozprzestrzenianych w powietrzu nawet poprzez przeszkody. Zmysł powonienia, a właściwie system VNO pozwala psom rozpoznać konkretną osobę lub zwierzę pośród wielu innych przedstawicieli tego samego gatunku. Świadczy to, że niezależnie od feromonów gatunkowych istnieją feromony osobnicze, różniące się prawdopodobnie składem ilościowym w ramach feromonu gatunkowego, a psy są w stanie wyczuć te subtelne różnice. Testy zapachowe zostały uznane jako dowody procesowe przez sądy, ponieważ substancje wydzielane przez poszczególnych ludzi są niepowtarzalne, pozostawiają osobnicze ślady, podobnie jak odciski palców. Przypuszcza się, że w przeszłości służyły one do rozpoznawania na odległość, tj. wyczuwania obecności osób najbliższych i rozróżniania ich od dalszych krewnych i obcych bez kontaktu wzrokowego, czy werbalnego. Ślad pozostawiony po określonej osobie mógł być rozpoznawalny przez długi okres nawet po jej odejściu. Obecnie możliwości korzystania z szóstego zmysłu zanikły prawie całkowicie u ludzi cywilizowanych, u psów pozostały, i podobnie jak każde inne zdolności percepcyjne mogą być poddawane doskonaleniu przez szkolenie. Możliwe, że niemowlęta, u których organ VNO jest silniej rozwinięty niż u ludzi dorosłych, są zdolne do rozpoznania matki i osób opiekującym się nimi na podstawie wydzielanych przez nie feromonów.
Na uwagę zasługują feromony służące do zaznaczania terytorium. Psy, wilki, czy lisy znaczą substancjami znajdującymi się w moczu terytorium, które uznają za teren swojego przebywania i polowania. W naturze wtargnięcie na terytorium zaznaczone przez osobnika tego samego lub zbliżonego gatunku oznacza inwazję i jest sygnałem do walki o obronę terytorium ze strony dotychczasowego użytkownika lub o zajęcie go z punktu widzenia intruza. U psów domowych zwyczaj zaznaczania terytorium, przez spryskanie, np. pnia drzewa odrobiną moczu, nawet na odległym od domu spacerze należy uznać za atawizm, ponieważ nie wywołuje to spodziewanych konsekwencji, natomiast przypomina pozostawiane często przez młodych ludzi napisy ''ja tu byłem''.
Feromony organizmów morskich
Ryby i inne zwierzęta żyjące w wodzie mają duże trudności w znalezieniu partnera płciowego, ponieważ woda jest środowiskiem gęstym, trudno przenikliwym dla większości sygnałów, zawiera zwykle dużo zanieczyszczeń chemicznych i mechanicznych (zawiesiny, mikroorganizmy), ponadto akweny wodne zajmują często duże przestrzenie. Z tej przyczyny feromony pełnią istotną funkcję w odnajdywaniu partnerów, synchronizacji cyklu rozrodczego i stymulowaniu zachowania się w trakcie tarła. Ponadto z uwagi na to, że wiele ryb i innych zwierząt wodnych, podobnie jak owady, tworzy kolonie nazywane ławicami, feromony umożliwiają im rozpoznawanie stopnia pokrewieństwa, a tym samym prawa do przebywania w grupie i udzielania sobie wzajemnej pomocy. U niektórych ryb w trakcie tarła dochodzi do wymiany feromonów pomiędzy partnerami w ściśle określonej kolejności. Każdy feromon wywołuję specyficzną reakcję i odpowiedź w postaci wydzielenia kolejnego feromonu przybliżającego finał. Zachowanie tych ryb przypomina wymianę miłosnych listów, rodzaj wyrafinowanego flirtu.
Zapłodnienie ikry w wodzie, tj. w przypadku, gdy nie dochodzi do zbliżenia fizycznego samic i samców, wymaga synchronizacji składania jaj i wydzielania mlecza. Woda szybko rozcieńcza nasienie, a jej ruch może oddalić od siebie gamety i skutecznie zapobiec ich połączeniu. Przebieg godów śledzia atlantyckiego (Clupea harengus pallasi) jest dobrym przykładem rozwiązania tego problemu przez naturę. Feromon indukujący składanie ikry przez samice jest wydzielany razem z mleczem. Działa on na dojrzałe seksualnie i przygotowane do rozrodu śledzie, zarówno na samice, jak i samce. Wprowadzenie tego feromonu, pod postacią świeżego śledziowego mlecza lub nawet roztworu po odwirowaniu z niego plemników, do akwarium z dojrzałymi do rozrodu śledziami, indukuje proces uwolnienia gamet, tzn. samice zaczynają składać ikrę, a samce wydzielać mlecz. Minimalne aktywne stężenie tego feromonu odpowiada rozcieńczeniu świeżego mlecza w stosunku 1:500. W odpowiedzi na feromon zawarty w mleczu, samice również wydzielają swoisty feromon o nieznanym dotychczas składzie. Głównymi składnikami feromonu występującego w spermie śledzia są wolne i związane steroidy. Ilościowy skład tego feromonu zmienia się w czasie sezonu rozrodczego. Wiele feromonów płciowych innych ryb okazało się także steroidami, analogami hormonów płciowych ssaków. Feromon płciowy złotej rybki (Carassium auratus) jest mieszaniną przynajmniej pięciu związków o budowie sterydowej. W tym feromonie znaleziono również prostaglandyny, np. PGF2<F128>a<F255>.
Tetrodokryna, znana jako jedna z najsilniejszych toksyn produkowanych przez organizmy morskie, służy puffer fish (Fugu niphobles) jako feromon powodujący wydzielanie mlecza przez samce w okresie składania ikry. Jest wytwarzana przez komórki jajowe samic i wydzielana bezpośrednio przez kloakę przed składaniem ikry. Minimalne stężenie aktywne tego feromonu wynosi 5 ng/l, tj. około 15 pM.
W czasie poprzedzającym składanie ikry, samice i samce Platynereis dumerilii i Nereis succinea gromadzą się w ławice. Gody rozpoczynają samce uwalniając feromon wywołujący składanie ikry przez samicę, po czym samice zaczynają pływać z dużą szybkością wewnątrz niedużego kręgu utworzonego przez samce. Po 10-40 sekundach takiego pływania samice zaczynają składać ikrę, razem z feromonem stymulującym samce do uwolnienia spermy. W wyniku spowodowanego przez pływające samice zawirowania wody wydzielone nasienie tworzy chmurę, która osadza się na ikrze.
Synchronizacja czasu składania jaj i wydzielania nasienia jest niezwykle ważna nie tylko dla ryb, ale i dla innych organizmów żyjących w wodzie, u których nie dochodzi do fizycznego kontaktu w trakcie zapładniania jaj. Składanie jaj do wody i wydzielanie spermy odbywać się musi nie tylko równocześnie, ale w tym samym miejscu, ponieważ woda, szczególnie będąca w ruchu, szybko rozcieńcza nasienie poniżej aktywnego stężenia. Często feromon wydzielany przez samce razem ze spermą indukuje nie tylko samice do składania jaj, ale równocześnie inne samce do wydzielania spermy. Również feromony wydzielane przez samice razem z jajami lub nawet bezpośrednio przed ich złożeniem wywołują u samców wydzielanie mlecza. Jest to tzw. efekt ''kuli śnieżnej'', powodujący masowe, wręcz epidemiczne wydzielanie gamet. Podobnym przykładem takiego postępowania jest zachowywanie się korali. Na ich dojrzewanie płciowe wpływa wiele czynników, w tym stopień naświetlenia, temperatura wody, fazy księżyca, a nawet czas pływów wywołujący potrzebny ruch wody. Natomiast sygnałem do masowego wydzielania gamet są męskie i żeńskie feromony.
Feromony węży
Feromony pełnią istotną rolę w rozmnażaniu węży. Znamiennym tego przykładem jest gatunek Thamnophilis sirtalis parietalis. Feromony płciowe znajdują się na skórze tych węży, a więc należą do feromonów powierzchniowych. Żeńskie atraktanty są mieszaniną długołańcuchowych (C29-C37) nasyconych i nienasyconych metyloketonów, podczas gdy męskie feromony zawierają skwalen i inne, jeszcze nie zidentyfikowane lipidy. W czasie godów tego gatunku, samce występujące w ogromnej przewadze liczbowej, 10-100/1 samicę, rozpoczynają zalotny bal tworząc kłębowisko kotłujących się ciał. Do nich dołączają samice. Poszczególne osobniki rozpoznają partnerów płciowych poprzez dotyk skóry językiem. Męskie feromony, jak się wydaje, służą jedynie jako ostrzeżenie w przypadku kontaktu z osobnikiem tej samej płci. Posmarowanie skóry samicy ekstraktem skóry samców powoduje, że nie jest przez samców traktowana jak samica. Natomiast zabieg przeciwny, tj. posmarowanie skóry samca ekstraktem skóry samic sprawia, że takie samce są traktowane jako samice, przy czym nie jest to objaw homoseksualizmu. Brak żeńskich feromonów na skórze samicy sprawia, że samce ją ignorują. Przypuszcza się, że zgromadzenie w jednym miejscu, w określonym czasie tysięcy węży następuje pod wpływem nie rozpoznanego dotychczas feromonu agregacyjnego.
Zastosowanie feromonów
Produkty spożywcze wytwarzane przez człowieka i przeznaczane na jego potrzeby są w znacznym stopniu niszczone, tzn. konsumowane przez mikroorganizmy, owady, czy gryzonie. Ludzie starają się zmniejszyć straty żywności, stosując między innymi insektycydy, które, niestety, często są toksyczne nie tylko dla owadów - szkodników, ale i dla owadów pożytecznych, a także dla organizmów wyższych, łącznie z ludźmi. Z tego powodu obserwuje się tendencję do zastępowania insektycydów innymi, bardziej selektywnymi środkami zwalczania szkodników. Dużą nadzieję w walce ze szkodnikami pokłada się w feromonach działających w niskich stężeniach, najczęściej nieszkodliwych dla większości organizmów. Mogą one zapewnić zwalczanie ściśle określonego gatunku owadów, w wybranej fazie rozwoju, np. poprzez zapobieganie zapłodnieniu, utrudnianie złożenia jaj, hamowanie przepoczwarzenia, itp. Specjalne pułapki na owady zawierające feromony umożliwiają odłowy określonych owadów w dowolnym czasie, korzystnie już na początku sezonu ich występowania, co zapewnia selektywne odłowy wybranych gatunków, nawet płci, i ich zwalczanie jeszcze przed stadium rozrodczym i intensywnym żerowaniem. Zastosowanie feromonów w zwalczaniu owadów ma przewagę nad innymi środkami przeciwowadowymi, ponieważ nie zatruwają one środowiska, nie wywołują efektu odporności u owadów, umożliwiają niszczenie szkodników przed stadium żerowania, a ich rejestracja, tj. zgoda na dopuszczenie do stosowania, odbywa się szybko i jest łatwiejsza do uzyskania niż dla pestycydów.
Feromonowe pułapki przeciwowadowe
W procesie zwalczania owadów feromony stosuje się najczęściej jako składniki pułapek. Feromony mają na celu zwabienie owadów do urządzeń uniemożliwiających ich opuszczenie lub powodujących fizyczne zniszczenie za pomocą środków toksycznych albo mechanicznych, np. taśm klejących. Pułapki przeciwowadowe były znane i stosowane od dawna, a jako przykład można podać lepy na muchy, które przyciągały owady zapachem lub smakiem, często miodowym czy też owocowym, zaś klejąca substancja pokrywająca lep krępowała ruchy owadów, uniemożliwiając im odlot. Ćmy łapano w nocy na podświetlonych ekranach. Pułapki feromonowe znacznie przewyższają skutecznością uprzednio wymienione, ponieważ owady przebywając nawet w dużej odległości od nich nie są w stanie oprzeć się przyciągającej sile wabików. Stężenia feromonów w pułapkach są na ogół większe niż w naturze, co powoduje zagłuszanie źródeł naturalnych, dezorientację odbiorców i kierowanie ich do pułapek. Właściwe dozowanie feromonów przez pułapkę decyduje o jej skuteczności. Urządzenie dozujące (dyspenser) powinno zapewniać określone, stałe stężenie feromonu w pobliżu pułapki oraz długi czas jej działania. Rodzaj dozownika dobiera się w zależności od lotności feromonu, wielkości pułapki i czasu jej działania. Do najczęściej stosowanych dozowników należą elementy o mikroporowatej strukturze, np. wiązki kapilar, włókna kapilarne, laminowane krążki, pakunki bibułowe, plastikowe porowate opaski lub ziarna, bądź pojemniki zamknięte porowatymi błonami o różnej przepuszczalności. Budowa pułapek zależy również od ich przeznaczenia, wielkości i czasu działania. Najprostsze przypominają znane lepy na muchy, a więc są taśmami lub opaskami pokrytymi substancjami klejącymi zawierającymi feromon. Bardziej skomplikowane są urządzeniami przestrzennymi (np. puszkami) o różnej pojemności z wejściem w kształcie lejka uniemożliwiającym wyjście, z wnętrzem pustym lub wypełnionym cieczą, a nawet insektycydami. Najczęściej stosowane pułapki feromonowe zawierają atraktanty płciowe lub feromony agregacyjne. Ostatnio coraz częściej używa się feromonów znaczących, których zadaniem jest dezinformacja, np. przekazanie samicom nieprawdziwej informacji o złożeniu już jaj na danym obiekcie (owocu, kwiecie, liściu itp.). Feromony takie mogą również przekazywać informacje o rzekomym zapłodnieniu samic.
Jak na razie feromony, mimo ogromnych oczekiwań, nie znalazły szerokiego zastosowania w ochronie roślin i produktów spożywczych. Głównymi przeszkodami w ich upowszechnieniu są wysokie ceny związane przede wszystkim z małym zapotrzebowaniem i brakiem właściwej promocji. Prawdopodobnie dużo wysiłku i czasu zajmie przekonanie producentów rolnych do zalet feromonów, jako skutecznych środków pielęgnacji upraw i lasów.
Aleksander Kołodziejczyk - rektor@pg.gda.pl
Politechnika Gdańska
Wydział Chemiczny
PPG 5/98
Praca wpłynęła do Chemfana: 27-08-99

