|
Paweł Łukasz Urban 35-125 Rzeszów ul. Solarza 22 m 13 plurban@friko5.onet.pl |
Paweł Mazur 41-300 Dąbrowa Górnicza ul. Augustynika 13a m 26 chemik@friko2.onet.pl |
| Rzeszów - Dąbrowa Górnicza, luty 1999 | |
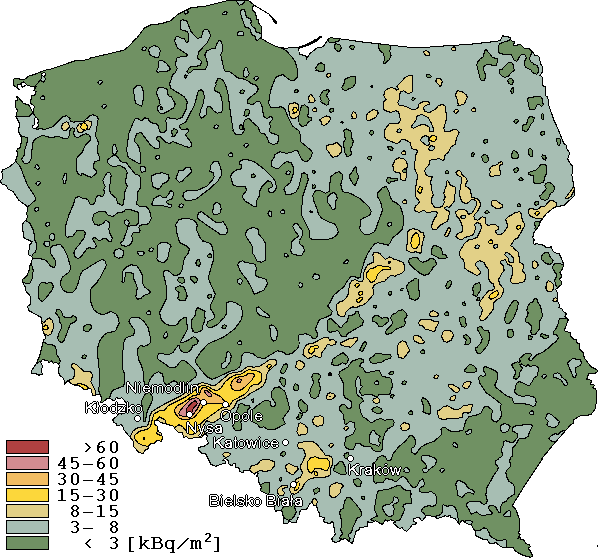
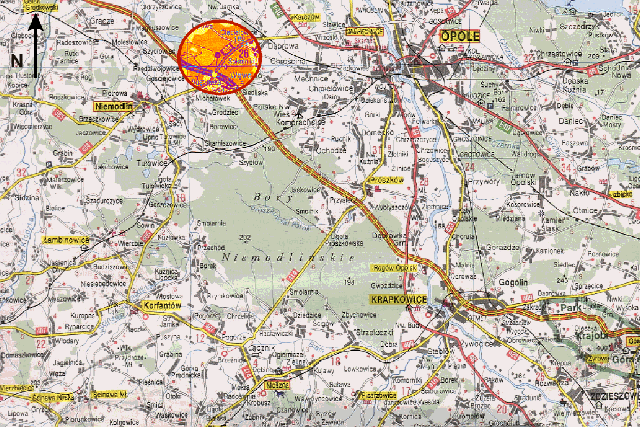
Rys.2 Mapa użytkowa okolic Niemodlina (PPWK, O12).
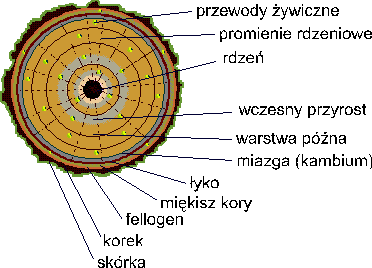
Rys.3 Przekrój pnia sosny.
Dalsza obróbka wiązała się ze zmniejszeniem objętości próbek,
a jednocześnie zwiększeniem koncentracji składników
nieorganicznych. W tym celu przeprowadzono spopielenie w
temperaturze 400 oC przez 6h. Jeżeli po takim zabiegu
w parowniczce pozostawał węgiel, kontynuowano spalanie przez
kolejną godzinę. Po spopieleniu próbki miały postać brązowego
proszku. Odnotowano masę próbek zarówno przed jak i po
spaleniu. Podczas obróbki wyników posługiwano się jednak masą
świeżych próbek.
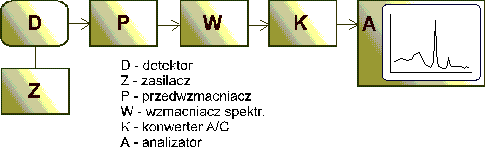
Należy zwrócić uwagę, że określenie aktywności 137Cs
sprowadza się do zmierzenia aktywności gamma baru 137mBa.
Cez-137 emituje cząstki beta-minus, rozpadając się na 137mBa.
Okres półrozpadu tego nuklidu wynosi 2,552 min. Rozpada się on
ze skończonym prawdopodobieństwem (89,98 %) na 137Ba
z wydzieleniem kwantu gamma o energii 661,65 keV - rejestrowanym
przez urządzenie. Potas 40K ulega przemianie beta,
spektrometr wykrywa kwanty gamma emitowane przez 40Ar
w stanie wzbudzonym (1460,8 keV). Prawdopodobieństwo rozpadu
nuklidu 40K wynosi 10,67 %.
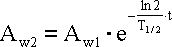
gdzie: Aw2 - aktualna aktywność próbki, Aw1
- aktywność oznaczona, T1/2 - czas półrozpadu
izotopu, t - czas pomiędzy oznaczeniem a pomiarami.
Aktywność każdej z próbek liczono na podstawie aktywności wzorca, z wzoru:
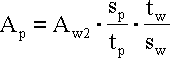
gdzie: Ap - aktywność próbki, sp -
powierzchnia pod krzywą Gaussa dopasowaną do piku nuklidu w
widmie próbki, tp - czas pomiaru próbki, sw
- powierzchnia pod krzywą Gaussa dla piku danego nuklidu w
widmie wzorca, tw - czas pomiaru wzorca.
Błąd pomiaru, wynikający z niedokładności rejestracji zliczeń w widmie próbki i wzorca policzono ze wzoru:
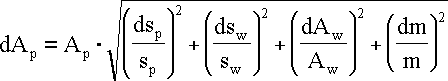
przy czym składnik odpowiadający względnemu błędowi pomiaru masy (dm/m) został zaniedbany ze względu na nieznaczne wartości.
Po zmierzeniu aktywności potasu-40 w próbkach możliwe staje się obliczenie wartości całkowitego stężenia potasu w drewnie. Wykorzystano tu wzór:
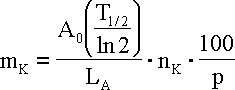
gdzie: mK - masa całkowita potasu, nK - masa molowa potasu promieniotwórczego (40 g/mol), p - ambudancja procentowa 40K w potasie (0,0117 %), T1/2 - czas półrozpadu 40K, LA - liczba Avogadro.
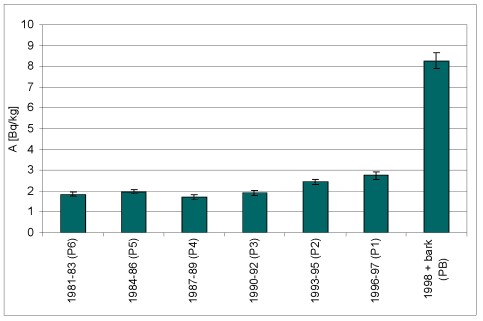
Wyk.1 Rejestracja zmian stężenia 137Cs
w próbkach kolejnych warstw przyrostów rocznych drewna i w
korowinie Pinus silvestris L.
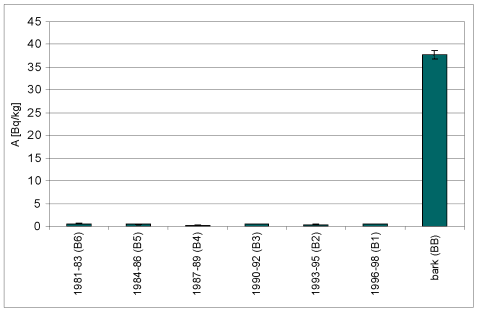
Wyk.2 Rejestracja zmian stężenia 137Cs
w próbkach kolejnych warstw przyrostów rocznych drewna i w
korowinie Betula pendula Roth.
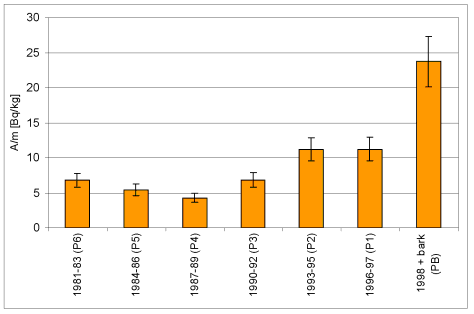
Wyk.3 Rejestracja zmian stężenia 40K
w próbkach kolejnych warstw przyrostów rocznych drewna i w
korowinie Pinus silvestris L.
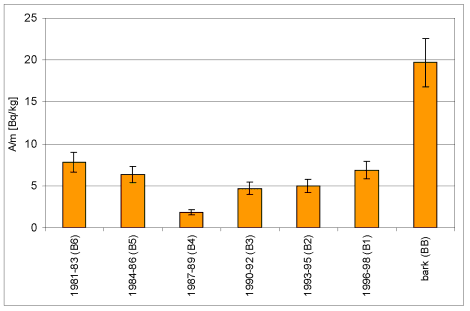
Wyk.4 Rejestracja zmian stężenia 40K
w próbkach kolejnych warstw przyrostów rocznych drewna i w
korowinie Betula pendula Roth.
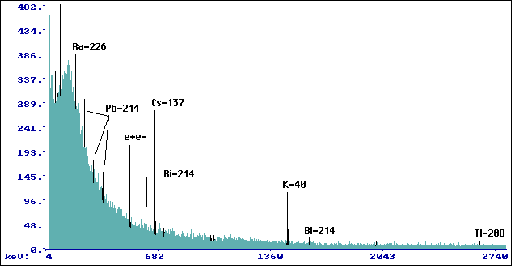
Wyk.5 Widmo dla warstwy korowej pnia Pinus
silvestris L. (tp=149880 s, mp=41,6 g).
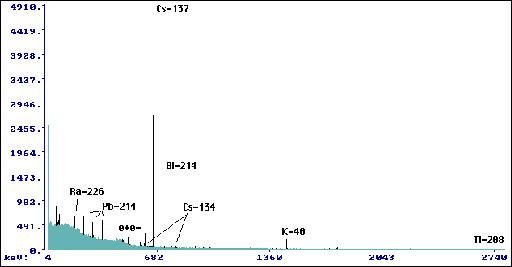
Wyk.6 Widmo dla warstwy korowej pnia Betula
pendula Roth. (tp=162240 s, mp=165,8 g).
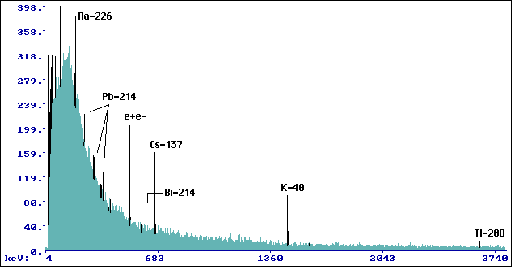
Wyk.7 Widmo dla przyrostów 1996-97 Pinus
silvestris L. (tp= 133200 s, mp=75,6 g).
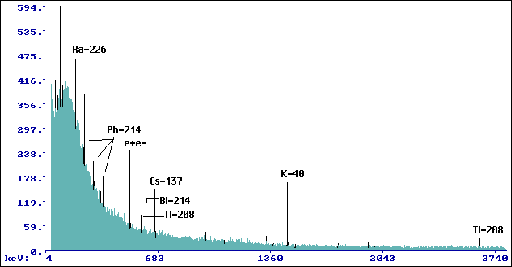
Wyk.8 Widmo dla przyrostów 1996-98 Betula
pendula Roth. (tp= 164520 s, mp=219,9 g).
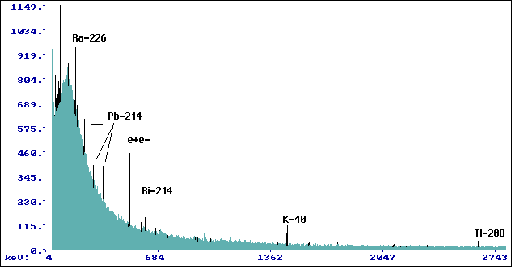
Wyk.9 Widmo dla promieniowania tła w domku
spektrometru (tp= 360000 s).
| Próbka | Wiek | m [g] | A [Bq] | A/m [Bq/kg] | +/-dA/m |
| P6 | 1981-83 | 97,5 | 0,18 | 1,84 | 0,10 |
| P5 | 1984-86 | 111,0 | 0,22 | 1,97 | 0,11 |
| P4 | 1987-89 | 101,2 | 0,17 | 1,70 | 0,12 |
| P3 | 1990-92 | 101,4 | 0,19 | 1,91 | 0,12 |
| P2 | 1993-95 | 101,4 | 0,25 | 2,44 | 0,13 |
| P1 | 1996-97 | 75,6 | 0,21 | 2,76 | 0,18 |
| PB | Kor. + 1998 | 41,6 | 0,34 | 8,27 | 0,40 |
Tab.2 Wyniki pomiaru 137Cs
w próbkach Betula pendula Roth.
| Próbka | Wiek | m [g] | A [Bq] | A/m [Bq/kg] | +/-dA/m |
| B6 | 1981-83 | 138,8 | 0,09 | 0,62 | 0,09 |
| B5 | 1984-86 | 130,9 | 0,06 | 0,48 | 0,07 |
| B4 | 1987-89 | 132,9 | 0,03 | 0,26 | 0,19 |
| B3 | 1990-92 | 155,9 | 0,08 | 0,53 | 0,07 |
| B2 | 1993-95 | 144,3 | 0,06 | 0,42 | 0,07 |
| B1 | 1996-98 | 219,9 | 0,12 | 0,54 | 0,05 |
| BB | Korowina | 165,8 | 6,26 | 37,8 | 0,89 |
Tab.3 Wyniki pomiaru 40K
w próbkach Pinus silvestris L.
| Próbka | Wiek | m [g] | A [Bq] | A/m [Bq/kg] | +/-dA/m |
| P6 | 1981-83 | 97,5 | 0,66 | 6,81 | 1,00 |
| P5 | 1984-86 | 111,0 | 0,61 | 5,47 | 0,82 |
| P4 | 1987-89 | 101,2 | 0,43 | 4,25 | 0,66 |
| P3 | 1990-92 | 101,4 | 0,69 | 6,80 | 1,04 |
| P2 | 1993-95 | 101,4 | 1,13 | 11,20 | 1,65 |
| P1 | 1996-97 | 75,6 | 0,85 | 11,20 | 1,71 |
| PB | kor. + 1998 | 41,6 | 0,99 | 23,70 | 3,56 |
Tab.4 Wyniki pomiaru 40K
w próbkach Betula pendula Roth.
| Próbka | Wiek | m [g] | A [Bq] | A/m [Bq/kg] | +/-dA/m |
| B6 | 1981-83 | 138,8 | 1,09 | 7,82 | 1,19 |
| B5 | 1984-86 | 130,9 | 0,83 | 6,37 | 0,96 |
| B4 | 1987-89 | 132,9 | 0,24 | 1,84 | 0,30 |
| B3 | 1990-92 | 155,9 | 0,74 | 4,73 | 0,71 |
| B2 | 1993-95 | 144,3 | 0,72 | 4,98 | 0,75 |
| B1 | 1996-98 | 219,9 | 1,51 | 6,86 | 1,01 |
| BB | Korowina | 165,8 | 3,27 | 19,70 | 2,85 |
Tab.5 Masa całkowita potasu [mikro-g,
ppm] w 1 g drewna Pinus silvestris L.
| Probka | Wiek | MK [mikro-g] | +/-dmK |
| P6 | 1981-83 | 225 | 33 |
| P5 | 1984-86 | 180 | 27 |
| P4 | 1987-89 | 140 | 22 |
| P3 | 1990-92 | 224 | 34 |
| P2 | 1993-95 | 369 | 55 |
| P1 | 1996-97 | 370 | 56 |
| PB | kor. + 1998 | 783 | 118 |
Tab.6 Masa całkowita potasu [mikro-g,
ppm] w 1 g drewna Betula pendula Roth.
| Próbka | Wiek | MK [mikro-g] | +/-dmK |
| B6 | 1981-83 | 258 | 39 |
| B5 | 1984-86 | 210 | 32 |
| B4 | 1987-89 | 61 | 10 |
| B3 | 1990-92 | 156 | 23 |
| B2 | 1993-95 | 164 | 25 |
| B1 | 1996-98 | 226 | 33 |
| BB | Korowina | 650 | 94 |
Praca wpłynęła na Chemfan: 15-04-99
